 2020, Vol. 32
2020, Vol. 32 


浮游植物作为水体中的重要初级生产者, 广泛存在于湖泊和河流等水体中, 在淡水系统的物质循环与能量流动中起着重要的作用[1-2].因其生长周期短、对环境变化较为敏感的特点, 浮游植物个体和群落被认为是指示环境变化的敏感指标[3].同时, 湖泊物理(如光照、温度)、化学(营养物、毒素)和生物作用(竞争、摄食、外来物种)因素等的改变均能够对浮游植物的群落组成和动态变化产生重要影响[4-8].随着流域开发的持续与人类活动的增强, 排放到湖泊中的营养物逐渐增多, 营养物排放负荷的累积导致水体富营养化, 出现部分浮游植物门类过量繁殖, 甚至导致群落优势种的明显转换, 水体环境质量和生态系统显著退化[9-11].
抚仙湖(24°17'~24°37'N, 102°49'~102°57'E)位于云贵高原西南部, 是珠江源头第一大湖和我国第二深水湖泊[12-13].抚仙湖作为典型的高原深水型淡水湖泊, 其蓄水量占云南省总蓄水量的78%, 是滇中地区社会经济可持续发展的重要资源保障[12-13].然而近年来, 随着人类活动的增强, 抚仙湖营养盐总体缓慢上升、水体透明度逐渐下降, 同时还面临外来物种入侵、气候变化等多种环境压力的胁迫, 湖泊生态系统功能呈退化趋势[14-19].同时, 抚仙湖流域的土地利用类型与开发强度存在较大的空间差异.其中, 湖盆北部流域主要包括城乡居民生活污染、农业开发、磷矿山和磷化工开发, 而湖盆南部流域主要为农业开发[20].流域开发的持续增强及其空间异质性可能对湖泊水体水质和生态系统结构产生影响.
抚仙湖浮游植物的调查研究始于1950s, 已有研究结果显示抚仙湖浮游植物的演变趋势明显.其中清洁性种类逐渐减少、喜营养种类增多, 转板藻逐渐占据优势, 同时藻类生物量和物种丰富度呈现增加的趋势[21-23]. 2002-2003年期间的抚仙湖浮游植物调查表明, 水体水温是驱动藻类群落逐月变化的主要环境因子, 且水体氮、磷等营养盐的季节变化特征不显著[23].然而在流域开发持续的背景下, 近年来抚仙湖水体化学特征的变化对水体硅藻群落产生了明显的驱动作用[24-25], 水体营养水平的持续上升已经对冬春季节浮游植物的演替产生了重要影响[26].总之, 抚仙湖已有研究集中于藻类生物量但缺乏群落结构变化的季节模式等周年调查[22-23, 27-30], 需要我们系统识别浮游植物群落变化的逐月模式、空间差异特征及其主要环境因子.通过认识抚仙湖生态系统关键组分的变化模式, 可对该湖泊开展生态安全的系统评价与有效的流域管理提供重要的科学依据.
本研究于2015年对抚仙湖浮游植物生物量、群落结构和优势种进行逐月调查, 运用方差分析(ANOVA)和群落相似性分析(ANOSIM)分别检验浮游植物生物量、环境因子和群落结构的时空差异, 结合主成分分析和断棍模型提取统计学上显著的主轴代表浮游植物群落结构变化, 通过多元统计分析(如方差分解)方法对影响浮游植物时空变化的显著环境因子进行识别, 以期为抚仙湖水环境保护及生态系统评价提供重要的数据支撑.
1 材料与方法 1.1 研究区概况抚仙湖位于云南省玉溪市澄江县、江川区、华宁县之间, 属南盘江水系, 是在云南高原第三纪抬升过程中所形成的构造断陷湖.受构造盆地影响, 其形态主要呈南北向分布, 为倒葫芦状, 两端大, 中间窄, 湖长约31.5 km, 最宽处约11.5 km, 湖岸线总长约88.2 km, 南部湖区较北部湖区窄且浅, 最大水深约155.0 m, 平均水深89.6 m, 相应湖面高程1721.0 m, 湖面面积211.0 km2, 流域面积1044.6 km2[12, 31].抚仙湖主要靠降雨和地表径流补给, 其入湖河流共计103条, 包括梁王河、东大河、路岐河、尖山河等, 多为间歇性河流.湖东岸的海口河为抚仙湖唯一的出水口, 出水量较小.抚仙湖湖区属于亚热带半湿润季风气候, 干湿季分明, 5-10月为雨季, 11-4月为旱季.
1.2 样品采集与处理根据抚仙湖形态特点设置南部湖区T1、中部湖区T2和北部湖区T3共3个采样点(图 1), 于2015年1-12月逐月采集表层水样.

|
图 1 抚仙湖采样点分布 Fig.1 Distribution of sampling sites in Lake Fuxian |
用5 L采水器采集表层水样(50 cm), 取1 L水样加入鲁哥和甲醛固定后避光带回实验室用于浮游植物鉴定.水样静置48 h, 吸去上层清液, 收集30 mL浓缩样品, 完全混合后取0.1 mL于容积为0.1 mL的计数框中, 在显微镜下计数, 每个样品计数两次, 若两次计数结果与其均数的差距大于15%, 则进行第3片计数.根据形态相似的几何学方法计算浮游植物细胞体积, 并转换为生物量结果[32], 浮游植物物种鉴定主要参照《中国淡水藻类——系统、分类及生态》[33].
取1 L表层水样用于水体理化指标的测定.主要方法为:参照《水与废水监测分析方法》(第四版)进行包括总氮(TN)和总磷(TP)浓度的测定[34].水温(WT)和pH采用多参数水质检测仪(美国YSI6600)现场测量, 透明度(SD)用塞氏盘现场测定.
1.3 数据统计分析本文中, 选择相对生物量大于10%的浮游植物属种为优势种[35].运用ANOSIM检验采样点浮游植物群落结构在时间和空间上的相异性, 相异系数R在0~1之间(如R值越低说明相异性越低)[36].进一步采用ANOVA检验3个采样点浮游植物生物量及各环境因子的时空差异.运用主成分分析(principal component analysis, PCA)、Pearson相关分析和方差分解(variance partitioning)识别影响浮游植物的主要环境因子.在探讨浮游植物群落结构与环境因子的关系时, 首先对浮游植物相对生物量数据进行平方根转换, 对转换后的数据进行去趋势对应分析(detrended correspondence analysis), 结果显示最大梯度长度为2.13 SD, 因此, 选择主成分分析进行排序分析, 并结合断棍模型(broken- stick)提取统计学上显著的主轴表征浮游植物群落结构变化.本文使用R3.2.2软件中的vegan、multcomp和psych软件包进行数据分析[37].
2 结果 2.1 主要环境因子抚仙湖2015年1-12月主要环境因子的变化范围及特征见图 2.采样期间, 水温在13.7~23.0℃之间, 最高水温23.0℃出现在8月, 最低水温13.7℃出现在2月.透明度在3.0~10.4 m之间, 1月最高, 为10.4 m, 3月最低, 为3.0 m. pH范围为8.0~9.0, 最高值出现在3月, 为9.0, 最低值出现在12月, 为8.0. TP浓度在10.0~65.0 μg/L之间, 在3月达到年内最高值, 为65.0 μg/L, 5月浓度最低, 3个点均 < 10.0 μg/L. TN浓度在130.0~360.0 μg/L之间, 8月浓度最大, 为360.0 μg/L, 11月浓度达到全年最低值130.0 μg/L.氮磷摩尔比(N:P)范围为2.9~36.0, 最高值出现在8月, 为36.0, 最低值出现在11月, 为2.9.

|
图 2 抚仙湖2015年主要环境因子的逐月变化 Fig.2 Temporal changes of major environmental factors of Lake Fuxian in 2015 |
对各环境因子的时空分布差异进行ANOVA分析, 结果显示各环境因子总体具有显著的时间差异(n=36, P < 0.05), 而空间差异不显著(n=36, P>0.05).
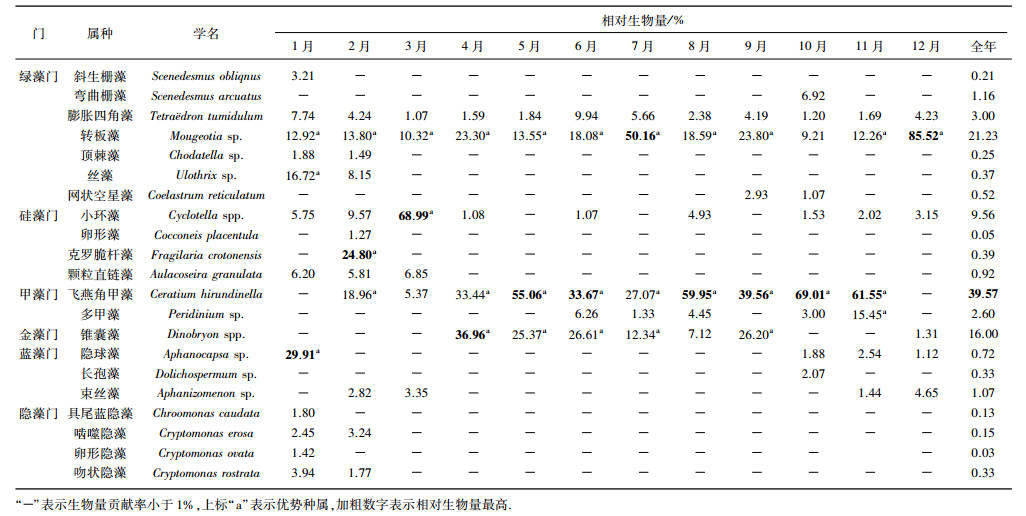
2.2 浮游植物的生物量、群落组成及优势种抚仙湖浮游植物由绿藻门、硅藻门、甲藻门、金藻门、蓝藻门和隐藻门组成.从相对生物量来看, 绿藻门比例全年较高; 硅藻门比例在早春和冬季比例较高, 1-3月份呈现逐渐上升的趋势; 甲藻门和金藻门分别在4-11月和4-9月比例较高; 蓝藻门1月比例相对较高; 隐藻门比例低于其他门类, 其冬季比例相对较高(图 3).从全年平均值来看, 绿藻门的转板藻(Mougeotia sp.)、甲藻门的飞燕角甲藻(Ceratium hirundinella)以及金藻门的锥囊藻(Dinobryon spp.)为优势种, 相对生物量均大于10%.从月平均来看, 绿藻门的转板藻为1-9月和11-12月的优势种, 且在7月和12月相对生物量最大, 10月其相对生物量为9.21%, 丝藻(Ulothrix sp.)为1月的优势种; 硅藻门的克罗脆杆藻(Fragilaria crotonensis)和小环藻(Cyclotella spp.)分别在2月和3月相对生物量最高; 甲藻门的飞燕角甲藻在2月和4-11月为优势种, 且在5-6月、8- 11月共6个月相对生物量最大, 多甲藻(Peridinium sp.)在11月百分比大于10%;金藻门的锥囊藻在4-7月和9、12月相对生物量大于10%, 且在4月相对生物量最高; 蓝藻门的隐球藻(Aphanocapsa sp.)1月相对生物量最大, 长孢藻(Dolichospermum sp.)、束丝藻(Aphanizomenon sp.)分别在10月、2-3月和11-12月相对生物量大于1%(表 1).

|
图 3 抚仙湖2015年浮游植物群落结构和生物量的逐月变化 Fig.3 Temporal changes of phytoplankton community structure and biomass in Lake Fuxian in 2015 |
| 表 1 抚仙湖相对生物量大于1%的浮游植物属种 Tab. 1 Phytoplankton taxa with relative biomass greater than 1% in Lake Fuxian |
抚仙湖浮游植物总生物量呈现双峰模式, 在4月和10月出现峰值(图 3).转板藻对绿藻门的贡献量最大, 其生物量4月和7月大于其他月份; 丝藻在1月对绿藻门贡献最大, 4月生物量最大; 克罗脆杆藻和小环藻的生物量和对硅藻门生物量的贡献分别于2月和3月达到最大; 飞燕角甲藻对甲藻门生物量的贡献最大, 其生物量在4月和10月出现双峰模式, 其中最大值出现在10月; 多甲藻对甲藻门生物量的贡献相对飞燕角甲藻较小, 其最大值出现在11月; 锥囊藻生物量在4-9月相对较高, 最大值出现在4月; 隐球藻1月对蓝藻门的贡献最大, 生物量最大值出现在10月(表 1, 图 4).

|
图 4 相对生物量大于10%的浮游植物种属生物量的逐月变化 Fig.4 Temporal biomass changes of phytoplankton taxa with relative biomass greater than 10% |
主成分分析结合断棍模型共提取到4个显著的主轴, 主轴1、2、3和4分别解释了浮游植物群落结构变化的43.60%、17.36%、10.70%和9.21%.抚仙湖浮游植物群落结构在属水平主要按照月份排序, 空间差异远小于时间差异. 4-11月样点主要位于轴1的正半轴, 甲藻门和金藻门占据主要优势, 12月、1-3月浮游植物主要位于轴1的负半轴, 蓝藻门1月比例较高, 硅藻门在2月和3月占据优势, 绿藻门在12月相对生物量最高(图 5).

|
图 5 抚仙湖浮游植物群落变化的主成分分析(□:T1;○:T2;△:T3.黑色:1月; 红色:2月; 蓝色: 3月; 绿色:4月; 深橙色:5月; 金色:6月; 紫红色: 7月; 暗紫色:8月; 深青色:9月; 红褐色: 10月; 玫瑰棕色:11月; 海绿色:12月) Fig.5 Principal component analysis of phytoplankton community changes in Lake Fuxian |
ANOSIM结果显示浮游植物群落结构存在显著的时间差异(R=0.7363, P < 0.001), 而空间差异不明显(R=-0.05841, P>0.05). ANOVA结果显示除隐藻门(时间及空间上均无显著差异)外, 浮游植物总生物量、各门类及优势种生物量在时间上差异显著(n=36, P < 0.001), 而在空间上无显著差异(n=36, P>0.05).
2.3 浮游植物与环境因子的关系相关分析结果显示(表 2), 浮游植物总生物量与水温呈显著正相关, 与透明度呈显著负相关; 隐球藻生物量与透明度呈显著正相关; 转板藻生物量与水温呈显著正相关; 丝藻生物量与水温呈显著负相关; 小环藻生物量与总磷浓度呈显著正相关, 与氮磷比和透明度呈显著负相关; 克罗脆杆藻生物量与总磷浓度呈显著正相关, 与氮磷比和水温呈显著负相关; 飞燕角甲藻生物量与水温呈显著正相关; 多甲藻生物量与水温呈显著正相关; 锥囊藻生物量与水温呈显著正相关.浮游植物群落PCA主轴1与水温呈显著正相关; 浮游植物群落PCA主轴2与总磷浓度呈显著负相关; 浮游植物群落PCA主轴3与透明度呈显著正相关.
| 表 2 抚仙湖浮游植物与环境因子的相关性矩阵 Tab. 2 Correlation matrix of phytoplankton and environmental factors in Lake Fuxian |
由于透明度与浮游植物存在着相互影响的关系, 因此进一步采用方差分解分析判断总磷浓度和水温对浮游植物群落变化的贡献量.方差分解结果(图 6)显示, 水温和总磷浓度共解释了抚仙湖浮游植物群落变化的31.6%, 其中水温和总磷浓度分别单独解释了浮游植物群落变化的26.0 %和2.6%, 两者共同解释了浮游植物群落变化的3.0%.

|
图 6 抚仙湖浮游植物群落结构与环境因子的方差分解 Fig.6 Variation partitioning results showing the explanatory powers of water temperature and total phosphorus concentration in accounting for phytoplankton community changes in Lake Fuxian |
2015年期间, 总磷是驱动抚仙湖浮游植物生物量和群落结构变化的重要环境因子之一.氮、磷是浮游植物生长的限制营养因子, 氮、磷营养元素的增加显著影响了湖泊浮游植物组成和分布, 促使水体浮游植物喜营养种类生物量显著增加, 而清洁型种类生物量显著下降甚至消失, 同时氮、磷营养盐也是驱动湖泊浮游植物季节变换的重要影响因子[38].本次调查表明, 总磷浓度与硅藻门的克罗脆杆藻和小环藻生物量呈显著正相关, 与代表浮游植物群落结构变化的主轴2呈显著负相关, 能够解释浮游植物群落结构变化的5.6%. 2015年1月上旬-3月中旬, 总磷浓度呈上升趋势, 2月和3月相对较高, 显著促进了克罗脆杆藻和小环藻的生长.总磷浓度对浮游植物产生显著影响, 也可能与抚仙湖在部分月份(1月、5-6月、8-9月)为磷限制状态有关(如较高的N:P比值).已有研究表明, 营养盐梯度也是驱动滇池、星云湖、洱海、阳宗海等大型高原湖泊浮游植物群落变化的重要环境因子[27, 39-43], 反映了区域社会经济发展导致的水体富营养化已成为影响高原湖泊藻类变化的重要驱动过程.
近年来抚仙湖由贫营养水体向贫-中营养水体过渡的趋势较为明显, 浮游植物群落结构及生物量的长期变化可能与水体营养水平升高有关.如与2002-2003年期间相比, 2015年抚仙湖平均总氮浓度由198.1 μg/L上升到231.1 μg/L, 平均总磷浓度由20.7 μg/L上升到23.0 μg/L.现代调查显示, 近几十年来抚仙湖浮游植物清洁型种类逐渐被喜营养种类取代. 1983年以来, 抚仙湖中转板藻逐渐占优且与营养水平上升的趋势同步, 同时银鱼引入导致的捕食压力变化也可能导致了大个体的转板藻占优[23-24, 26]. 2002年9月-2003年8月的逐月调查表明, 绿藻门的转板藻为全年的优势种[22-23], 而2015年期间锥囊藻和飞燕角甲藻的优势显著增加, 与转板藻共同成为主要的优势种且飞燕角甲藻年平均相对生物量均大于转板藻.同时转板藻、飞燕角甲藻和锥囊藻生物量均出现显著增长且抚仙湖浮游植物平均总生物量由2002-2003年的0.419 mg/L上升到2015年1-12月0.782 mg/L, 与沉积物记录的湖泊初级生产力的升高相一致[44].已有研究表明, 转板藻适合在贫中营养水体中生存, 飞燕角甲藻常见于中富营养、超富营养水体, 锥囊藻通常生长在贫-中营养水体[45].
抚仙湖浮游植物群落结构与云南高原其他深水湖泊相比差异较大, 这可能与营养水平等生境状态的不同有关.如贫营养湖泊泸沽湖浮游植物物种组成多样, 以硅藻门和绿藻门为主; 在中营养水平的阳宗海中, 蓝藻门为绝对优势种, 其中伪鱼腥藻、浮丝藻、束丝藻为全年的优势种.而在富营养化的程海中, 蓝藻门为优势门类, 包括卷曲鱼腥藻、固氮鱼腥藻、铜绿微囊藻、水华微囊藻和水华束丝藻[46-48].
3.2 温度和气候特征对浮游植物时空变化的影响模式温度是影响浮游植物生长的关键因子, 在浮游植物生物量和群落结构的变化过程中起到了重要的作用[49].本次调查中, 水温是驱动抚仙湖2015年浮游植物生物量和群落结构变化的显著环境因子, 能够解释群落结构变化的29.0%.温度通过影响新陈代谢而直接影响浮游植物的生长, 不同种类浮游植物对温度的响应存在差别[50].如硅藻相对喜低温, 由于硅质外壳较重及缺乏鞭毛而容易在水体中沉降, 因此在湖泊水体混合期间容易占据优势.本研究中, 硅藻门生物量和比例在1-3月和12月相对较高, 克罗脆杆藻和小环藻相对生物量分别在2月和3月最大.克罗脆杆藻为喜冷水种, 本研究中克罗脆杆藻生物量与水温呈显著负相关, 2015年2月水温最低(13.8℃), 其生物量最高, 与2002-2003年12月最高有所不同.
此外, 温度能够通过影响湖泊热力学分层而对深水湖泊浮游植物群落的构建及分布产生间接影响[51].抚仙湖为单次混合型湖泊, 1-3月和12月抚仙湖水温相对较低(水温范围为13.7~17.4℃), 湖泊0~50 m范围内水体基本处于混合状态. 4-11月, 抚仙湖水温相对较高(水温范围为17.1~23.0 ℃), 湖泊处于分层期[25].飞燕角甲藻通常对水体混合和低光照较敏感, 水体分层期生物量一般较高, 常在夏季浮游植物群落中占优势[45, 52], 同时早夏是锥囊藻最适宜其生长的最适时期[45, 53].本次调查表明, 抚仙湖水温的升高有利于易在分层期获得最佳生长的藻类(如飞燕角甲藻和锥囊藻)生长.如在湖水分层期间, 飞燕角甲藻生物量和相对生物量均较高, 为2月、4-11月的优势种且在5-6月和8-11月期间相对生物量最高.而锥囊藻为4-7月和9月的优势种, 4月生物量(2.23 mg/L)和相对生物量均最高.飞燕角甲藻和锥囊藻生物量均与水温呈显著正相关.
全球气候变暖的背景下, 温度升高能够加强湖泊热力学分层而减弱水体垂直混合, 使热力学分层期延长, 从而导致了个体小、沉降速率低的藻类丰度增加[54-55].此外, 温度升高能够延长藻类的最适生长期的时间[54]. 2002-2015年间, 抚仙湖流域年平均气温呈现波动上升趋势, 由16.2℃上升到17.1℃(红塔气象站), 而年平均水温则同步由18.2℃升高到18.9℃[56].本次调查中, 沉降速率较低、个体较小的硅藻小环藻在1-12月均有分布, 除5月、7月和9月外, 其余月份小环藻相对生物量均大于1%;而2002-2003年的调查显示, 小环藻仅在1-4月、11-12月出现. 2002-2003年期间, 锥囊藻出现在6-11月, 9月生物量最高[23]; 2005年6-7月锥囊藻为优势种[39]; 2010年9月锥囊藻在湖心区成为明显的优势种[57]; 2014年7月锥囊藻为优势种之一[58].而本研究中, 锥囊藻在4-12月均有出现.转板藻生存范围较广, 由于具有良好的光捕获能力和中等的沉降速率, 其既能耐受冬季水体混合和低光照, 也能在水体分层期获得最大生物量[45, 59-60].本研究中, 除10月相对生物量略低于10%外, 其余月份转板藻均为优势种, 其生物量与水温呈显著正相关. 2015年转板藻生物量于湖水分层期达到最高值且在12月相对比例最高, 与2002-2003年12月生物量和相对生物量均为最高有所不同.
2015年期间, 抚仙湖浮游植物群落结构和生物量均具有显著的逐月变化特征, 而空间差异不显著, 反映了环境因子季节变化的驱动作用.本次调查结果与李蕊等识别的硅藻群落结构时空分布特征相似[25], 而与潘继征等[30]、此里能布等[39]、蒋伊能等[26]和秦洁等[61]的结果不一致.这可能与后者包含了较多位于沿岸带的采样点有关, 而本研究和李蕊等[25]所选取的采样点均分布在远离湖滨带的敞水区.此外, 在抚仙湖盛行风的持续影响下浮游植物扩散能力更强也可能导致敞水区浮游植物空间分布的同质性增强[25, 30].
4 结论本次抚仙湖的逐月调查结果显示, 温度和总磷浓度是影响浮游植物生物量和群落结构季节变化的显著因子, 且水体环境与藻类群落分布的空间差异模式不显著.与2002-2003年绿藻门的转板藻生物量在全年占优相比, 2015年甲藻门的飞燕角甲藻和金藻门的锥囊藻生物量和相对生物量显著增加, 与转板藻共同成为主要优势种, 且飞燕角甲藻年平均相对生物量大于转板藻.近年来, 抚仙湖水体营养盐浓度的升高可能是导致浮游植物生物量和优势种变化的重要因素.因此, 在区域气候变化和流域开发持续的背景下, 控制流域污染物的输入并应对气候变暖是维持抚仙湖浮游植物群落结构和生态系统安全的重要前提.
致谢:感谢云南省高原地理过程与环境变化重点实验室各位老师和同学在野外采样和室内分析中给予的帮助.
| [1] |
Liu JK ed. Advanced aquatic biology. Beijing: Science Press, 1999. [刘建康.高级水生生物学.北京: 科学出版社, 1999. ]
|
| [2] |
Field CB. Primary production of the biosphere:Integrating terrestrial and oceanic components. Science, 1998, 281(5374): 237-240. DOI:10.1126/science.281.5374.237 |
| [3] |
Doney SC, Ruckelshaus M, Emmett Duffy J et al. Climate change impacts on marine ecosystems. Annual Review of Marine Science, 2012, 4(1): 11-37. DOI:10.1146/annurev-marine-041911-111611 |
| [4] |
Suikkanen S, Laamanen M, Huttunen M. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea. Estuarine, Coastal and Shelf Science, 2007, 71(3/4): 580-592. DOI:10.1016/j.ecss.2006.09.004 |
| [5] |
Tian C, Pei HY, Hu WR et al. Phytoplankton variation and its relationship with the environmental factors in Nansi Lake, China. Environmental Monitoring and Assessment, 2013, 185(1): 295-310. DOI:10.1007/s10661-012-2554-8 |
| [6] |
Chen M, Fan M, Liu R et al. The dynamics of temperature and light on the growth of phytoplankton. Journal of Theoretical Biology, 2015, 385: 8-19. DOI:10.1016/j.jtbi.2015.07.039 |
| [7] |
Jiang YJ, He W, Liu WX et al. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu). Ecological Indicators, 2014, 40: 58-67. DOI:10.1016/j.ecolind.2014.01.006 |
| [8] |
Leland HV. The influence of water depth and flow regime on phytoplankton biomass and community structure in a shallow, lowland river. Hydrobiologia, 2003, 506. DOI:10.1023/b:hydr.0000008596.00382.56 |
| [9] |
Wan L. Rules of phytoplankton dominant species formation in eutrophic water[Dissertation]. Nanjing:Hohai University, 2008. [万蕾.富营养化水体中浮游植物优势种形成规律研究[学位论文].南京: 河海大学, 2008. ]
|
| [10] |
Li ZJ, Chen XL, Zheng HL et al eds. Ecology. Beijing: Science Press, 2014: 156. [李振基, 陈小麟, 郑海雷等.生态学.北京: 科学出版社, 2014: 156. ]
|
| [11] |
Wang JL ed. Environmental microbiology. Beijing: Higher Education Press, 2004. [王家玲.环境微生物学.北京: 高等教育出版社, 2004. ]
|
| [12] |
Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences ed. Fuxian Lake. Beijing: Ocean Press, 1990. [中国科学院南京地理与湖泊研究所.抚仙湖.北京: 海洋出版社, 1990. ]
|
| [13] |
Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences ed. Report on lake survey in China. Beijing: Science Press, 2019. [中国科学院南京地理与湖泊研究所.中国湖泊调查报告.北京: 科学出版社, 2019. ]
|
| [14] |
Gao W, Chen Y, Xu M et al. Trend and driving factors of water quality change in Lake Fuxian (1980-2011). J Lake Sci, 2013, 25(5): 635-642. [高伟, 陈岩, 徐敏等. 抚仙湖水质变化(1980-2011年)趋势与驱动力分析. 湖泊科学, 2013, 25(5): 635-642. DOI:10.18307/2013.0503] |
| [15] |
Qin JH. Ecological effects of the invasive icefish Neosalanx taihuensis in a deep oligotrophic plateau lake, Lake Fuxian[Dissertation]. Wuhan:Institute of Hydrobiology, Chinese Academy of Science, 2005:20-25. [覃剑晖.太湖新银鱼入侵高原深水贫营养湖泊——抚仙湖的生态学效应[学位论文].武汉: 中国科学院水生生物研究所, 2005: 20-25. ]
|
| [16] |
Li YL, Gong ZJ, Shen J. Effects of eutrophication and temperature on Cyclotella rhomboideo-elliptica Skuja, endemic diatom to China. Phycological Research, 2012, 60(4): 288-296. DOI:10.1111/j.1440-1835.2012.00659.x |
| [17] |
Dong J, Zhou QC, Gao YN et al. Long-term effects of temperature and nutrient concentrations on the phytoplankton biomass in three lakes with differing trophic statuses on the Yungui Plateau, China. Annales De Limnologie-International Journal of Limnology, 2018, 54: 9. DOI:10.1051/limn/2017031 |
| [18] |
Zhou QC, Zhang YL, Li KD et al. Seasonal and spatial distributions of euphotic zone and long-term variations in water transparency in a clear oligotrophic Lake Fuxian, China. Journal of Environmental Sciences, 2018, 72: 185-197. DOI:10.1016/j.jes.2018.01.005 |
| [19] |
Zhou QC, Wang WL, Huang LC et al. Spatial and temporal variability in water transparency in Yunnan Plateau lakes, China. Aquatic Sciences, 2019, 81(2): 36. DOI:10.1007/s00027-019-0632-5 |
| [20] |
Yuxi Municipal People's Government ed. The 12th five-year plan for comprehensive prevention and control of water pollution in Fuxian lake basin. Beijing: Chinese Society of Environmental Sciences, 2012. [玉溪市人民政府.抚仙湖流域水污染综合防治"十二五"规划.北京: 中国环境科学学会, 2012. ]
|
| [21] |
Li YX, Wang L, Qi YK et al. Studies on the phytoplankton development trend in Lake Fuxian, China. J Lake Sci, 2007, 19(2): 223-226. [李荫玺, 王林, 祁云宽等. 抚仙湖浮游植物发展趋势分析. 湖泊科学, 2007, 19(2): 223-226. DOI:10.18307/2007.0218] |
| [22] |
Zhang X, Xie P, Chen FZ et al. Present status and changes of the phytoplankton community after invasion of Neosalanx taihuensis since 1982 in a deep oligotrophic plateau lake, Lake Fuxian in the subtropical China. Journal of Environmental Sciences, 2005, 17(3): 389-394. |
| [23] |
Zhang X, Xie P, Chen FZ et al. Driving forces shaping phytoplankton assemblages in two subtropical plateau lakes with contrasting trophic status. Freshwater Biology, 2007, 52(8): 1463-1475. DOI:10.1111/j.1365-2427.2007.01776.x |
| [24] |
Chen XL, Chen GJ, Lu HB et al. Long-term diatom biodiversity responses to productivity in lakes of Fuxian and Dianchi. Biodiversity Science, 2015, 23(1): 89-100. [陈小林, 陈光杰, 卢慧斌等. 抚仙湖和滇池硅藻生物多样性与生产力关系的时间格局. 生物多样性, 2015, 23(1): 89-100.] |
| [25] |
Li R, Chen GJ, Kang WG et al. Spatio-temporal variations of diatom community and their relationship with water environment in Fuxian lake. Environmental Science, 2018, 39(7): 3168-3178. [李蕊, 陈光杰, 康文刚等. 抚仙湖硅藻群落的时空变化特征及其与水环境的关系. 环境科学, 2018, 39(7): 3168-3178. DOI:10.13227/j.hjkx.201710188] |
| [26] |
Jiang YN, Zhang T, Chen L et al. Ldentification of phytoplankton community changes and environmental factors during the winter-spring transition in Fuxian Lake. Transactions of Oceanology and Limnology, 2016(6): 87-98. [蒋伊能, 张涛, 陈丽等. 抚仙湖冬-春季浮游植物群落结构变化及其影响因子识别. 海洋湖沼通报, 2016(6): 87-98.] |
| [27] |
Li SH, Yu MJ, Li GZ et al. Limnological survey of the lakes of Yunnan Plateau. Oceanologia et Limnologia Sinica, 1963, 5(2): 87-113. [黎尚豪, 俞敏娟, 李光正等. 云南高原湖泊调查. 海洋与湖沼, 1963, 5(2): 87-113.] |
| [28] |
Wei YX, Shi ZX et al eds. A survey of the phytoplankton and assessment of the water quality and tropic characteristics of three big lakes from Yunnan plateau//Compilation of reports on the survey of algal resources in south-western China. Beijing: Science Press, 1994. [魏印心, 施之新等.云南高原三大湖泊浮游植物调查及水质和热带特征评估//中国西南地区藻类资源调查报告的汇编.北京: 科学出版社, 1994. ]
|
| [29] |
Deng XY, Xu JH. Studies on algae of Fu-Xian Lake in Cheng-Jiang County, Yunnan. Journal of Yunnan University:Natural Sciences, 1996, 18(2): 139-145. [邓新晏, 许继宏. 澄江抚仙湖藻类植物研究. 云南大学学报:自然科学版, 1996, 18(2): 139-145.] |
| [30] |
Pan JZ, Xiong F, Li WC et al. Structure, distribution and its impact factors of phytoplankton community in Fuxian Lake. Acta Ecologica Sinica, 2009, 29(10): 5376-5385. [潘继征, 熊飞, 李文朝等. 抚仙湖浮游植物群落结构、分布及其影响因子. 生态学报, 2009, 29(10): 5376-5385. DOI:10.3321/j.issn:1000-0933.2009.10.024] |
| [31] |
Wang SM, Dou HS eds. Records of lakes in China. Beijing: Science Press, 1998. [王苏民, 窦鸿声.中国湖泊志.北京: 科学出版社, 1998. ]
|
| [32] |
Sun J, Liu DY, Qian SB. Study on phytoplankton biomass. Acta Oceanologica Sinica, 1999, 21(2): 75-85. [孙军, 刘冬艳, 钱树本. 浮游植物生物量研究. 海洋学报, 1999, 21(2): 75-85. DOI:10.3321/j.issn:0253-4193.1999.02.010] |
| [33] |
Hu HJ, Wei YX eds. Chinese freshwater algae-System, classification and ecology. Beijing: Science Press, 2006. [胡鸿钧, 魏印心.中国淡水藻类——系统、分类及生态.北京: 科学出版社, 2006. ]
|
| [34] |
Editorial board of "monitoring and analysis methods of water and wastewater", State Environmental Protection Administration of China ed. Minoring and analysis methods of water and wastewater: fourth edition. Beijing: China Environmental Science Press, 2002: 243-285. [国家环境环保总局《水和废水监测分析方法》编委会.水和废水监测分析方法: 第4版.北京: 中国环境科学出版社, 2002: 243-285. ]
|
| [35] |
Wu ZS, Cai YJ, Liu X et al. Temporal and spatial variability of phytoplankton in Lake Poyang:The largest freshwater lake in China. Journal of Great Lakes Research, 2013, 39(3): 476-483. DOI:10.1016/j.jglr.2013.06.008 |
| [36] |
Chen GJ, Dalton C, Leira M et al. Diatom-based total phosphorus (TP) and pH transfer functions for the Irish Ecoregion. Journal of Paleolimnology, 2008, 40(1): 143-163. DOI:10.1007/s10933-007-9148-4 |
| [37] |
Borcard D, Gillet F, Legendre P eds. Translated by Lai JS. Quantitative ecology-Application of R language. Beijing: Higher Education Press, 2014. [Borcard D, Gillet F, Legendre P著.赖江山译.数量生态学——R语言的应用.北京: 高等教育出版社, 2014. ]
|
| [38] |
Katherine RM, Rochelle GL, Mike C et al. Phosphorus availability, phytoplankton community dynamics and taxon-specific phosphorus status in the Gulf of Aqaba, Red Sea. Limnology and Oceanography:Methods, 2007, 52(2): 873-885. DOI:10.4319/lo.2007.52.2.0873 |
| [39] |
Cili nengbu, Mao JZ, Zhao HG et al. The phytoplankton community structure in Fuxian lake in the autumn of 2011. Environmental Science Survey, 2013, 32(3): 26-30. [此里能布, 毛建忠, 赵华刚等. 2011年抚仙湖浮游植物群落结构研究. 环境科学导刊, 2013, 32(3): 26-30. DOI:10.3969/j.issn.1673-9655.2013.03.007] |
| [40] |
Wang Z, Wang YC, Hu MM et al. Succession of the phytoplankton community in response to environmental factors in north Lake Erhai during 2009-2010. Fresenius Environmental Bulletin, 2011, 20(9): 2221-2231. |
| [41] |
Dai GY, Li J, Li L et al. The spatio-temporal pattern of phytoplankton in the north basin of Lake Dianchi and related environmental factors. Acta Hydrobiologica Sinica, 2012, 36(5): 946-956. [代龚圆, 李杰, 李林等. 滇池北部湖区浮游植物时空格局及相关环境因子. 水生生物学报, 2012, 36(5): 946-956.] |
| [42] |
Wang H, Yang SP, Fang SZ et al. Canonical correspondence analysis of relationship between characteristics of phytoplankton community and environmental factors in Dianchi Lake. China Environmental Science, 2016, 36(2): 544-552. [王华, 杨树平, 房晟忠等. 滇池浮游植物群落特征及与环境因子的典范对应分析. 中国环境科学, 2016, 36(2): 544-552. DOI:10.3969/j.issn.1000-6923.2016.02.034] |
| [43] |
Huang J. Studies on diversity of phytoplankton in the north of Lake Erhai and the taxonomy of Psephonema aenigmaticum (Chlorophyta)[Dissertation]. Nanchang: Jiangxi Normal University, 2014. [黄静.洱海北部湖湾浮游植物多样性及暗丝藻分类学研究[学位论文].南昌: 江西师范大学, 2014. ]
|
| [44] |
Shi HB, Chen GJ, Lu HB et al. Regional pattern of Bosmina responses to fish introduction and eutrophication in four large lakes from Southwest China. Journal of Plankton Research, 2016, 38(3): 443-455. DOI:10.1093/plankt/fbv118 |
| [45] |
Reynolds CS, Huszar V, Kruk C et al. Towards a functional classification of the freshwater phytoplankton. Journal of Plankton Research, 2002, 24(5): 417-428. DOI:10.1093/plankt/24.5.417 |
| [46] |
Wu JL, Jiang XZ, Xia WL et al. Climate and primary productivity over the last 500 years of the Chenghai Lake, Yunnan. Marine Geology & Quaternary Geology, 2002, 22(2): 95-98. [吴敬禄, 蒋雪中, 夏威岚等. 云南程海近500年来湖泊初始生产力的演化. 海洋地质与第四纪地质, 2002, 22(2): 95-98.] |
| [47] |
Dong YX, Tan ZW, Guo YY. The preliminary study on phytoplankton in Lugu Lake. Journal of Hydroecology, 2012, 33(3): 46-52. [董云仙, 谭志卫, 郭艳英. 泸沽湖浮游植物的初步研究. 水生态学杂志, 2012, 33(3): 46-52.] |
| [48] |
Bai NJ, Chen L, Jiang YN et al. Spatio-temporal characteristics of phytoplankton distribution and the identification of driving factors in the arsenic-contaminated Yangzong Lake. J Lake Sci, 2019, 31(1): 147-158. [白宁静, 陈丽, 蒋伊能等. 阳宗海砷污染背景下浮游植物的时空分布特征及其驱动因子解析. 湖泊科学, 2019, 31(1): 147-158. DOI:10.18307/2019.0114] |
| [49] |
Rhee GY, Gotham IJ. The effect of environmental factors on phytoplankton growth:Temperature and the interactions of temperature with nutrient limitation. Limnology and Oceanography, 1981, 26(4): 635-648. DOI:10.4319/lo.1981.26.4.0635 |
| [50] |
Lehman PW. The influence of climate on phytoplankton community biomass in San Francisco Bay Estuary. Limnology and Oceanography, 2000, 45(3): 580-590. DOI:10.4319/lo.2000.45.3.0580 |
| [51] |
Deng JM, Qin BQ. A review on studies of effects of climate change on phytoplankton in freshwater systems. J Lake Sci, 2015, 27(1): 1-10. [邓建明, 秦伯强. 全球变暖对淡水湖泊浮游植物影响研究进展. 湖泊科学, 2015, 27(1): 1-10. DOI:10.18307/2015.0101] |
| [52] |
Reynolds CS. Notes on the phytoplankton periodicity of Rostherne mere, Cheshire, 1967-1977. British Phycological Journal, 1978, 13(4): 329-335. DOI:10.1080/00071617800650391 |
| [53] |
Butterwick C, Heaney SI, Talling JF. Diversity in the influence of temperature on the growth rates of freshwater algae, and its ecological relevance. Freshwater Biology, 2004, 50(2): 291-300. DOI:10.1111/j.1365-2427.2004.01317.x |
| [54] |
Paerl HW, Huisman J. CLIMATE:blooms like it hot. Science, 2008, 320(5872): 57-58. DOI:10.1126/science.1155398 |
| [55] |
Winder M, Reuter JE, Schladow SG. Lake warming favors small-sized planktonic diatom species. Proceedings of the Royal Society B:Biological Sciences, 2009, 276(1656): 427-435. DOI:10.1098/rspb.2008.1200 |
| [56] |
Gu GH. Analysis of Fuxianhu lake water temperature features and change trend. Pearl River, 2008, 29(5): 38-39, 57. [谷桂华. 抚仙湖水温特征及趋势分析. 人民珠江, 2008, 29(5): 38-39, 57. DOI:10.3969/j.issn.1001-9235.2008.05.012] |
| [57] |
Qiu JR, Yang JX, Xia LL et al. Study on the aquatic ecosystem of Fuxian Lake. Ecological Economy, 2011(2): 379-383, 393. [邱家荣, 扬君兴, 夏黎亮等. 抚仙湖水生生态研究. 生态经济:学术版, 2011(2): 379-383, 393.] |
| [58] |
Niu Y, Kong XH, Yu H et al. Spatial distribution of phytoplankton community during summer stratification in Lake Fuxian. Chinese Journal of Ecology, 2016, 35(7): 1865-1871. [牛远, 孔祥虹, 余辉等. 抚仙湖夏季热分层时期浮游植物空间分布特征. 生态学杂志, 2016, 35(7): 1865-1871. DOI:10.13292/j.1000-4890.201607.019] |
| [59] |
Reynolds CS ed. Ecology of phytoplankton. Cambridge: Cambridge University Press, 2006.
|
| [60] |
Tapolczai K, Anneville O, Padisák J et al. Occurrence and mass development of Mougeotia spp. (Zygnemataceae) in large, deep lakes. Hydrobiologia, 2015, 745(1): 17-29. DOI:10.1007/s10750-014-2086-z |
| [61] |
Qin J, Wu XH, Wang Q et al. Community structure and biodiversity of phytoplankton in Fuxian Lake. Journal of Hydroecology, 2016, 37(5): 15-22. [秦洁, 吴献花, 王泉等. 抚仙湖浮游植物群落结构特征及多样性研究. 水生态学杂志, 2016, 37(5): 15-22. DOI:10.15928/j.1674-3075.2016.05.003] |